


A Simple Biophysical/Economic Model Explaining ‘Cultural’ Shifts between Cooperation and Competition, Comity and Discord
The root source of modern human discord re-identified from 'basic principles'
Introduction
The last Grundvilk post briefly discussed a few characteristics of mutualism; i.e., the mutually-beneficial relationships that sometimes exist between categorically different organisms, or groups of organisms. Through time and differing circumstance, for example, human beings with different strengths and weaknesses have frequently found ways to simultaneously benefit both themselves and others through cooperation. For much of human history, in fact, the net benefits received by these interacting individuals and/or groups have materially exceeded those obtainable by independent action.
Like in so many other things in nature, however, diverse qualities of mutualism exist. At the one extreme, groups, species, or individual organisms bound strongly together by obligate mutualism could not exist at all except for the sustained operation of that close mutualistic arrangement. Typically, this kind of mutualism exists when the mutualism participants dwell in a very resource-poor environment, and/or when the competencies (e.g., genotypes and/or phenotypes) of the participants concerned are specialized or otherwise restricted.1
All mutualist interactions come at some cost to the participants. If this cost is less than the benefit received from the mutualist arrangement, the arrangement will tend to continue. In the case of obligate mutualism, if costs of participating in that form of mutualism increase because of a change in the behavior or numbers of a partner mutualist, or because of a change in the external environment, then an obligate mutualist moiety will become unstable and can ultimately disappear. If one or both obligate mutualist partners can obtain needed resources directly from the environment, or from alternative partners, then obligate mutualism can transform into weaker facultative mutualism. At the extreme, if deleterious changes in partner or environment are much too abrupt to adjust to in a timely fashion, it is possible that members of the obligate mutualist ‘society’ will become at least locally extinct.
In parasitism and competition (including endpoint competitive exclusion), species and groups move from mutualist cooperation with others to parasitism and/or competition with others. In such relations, a species or group with some sort of temporary or permanent absolute advantage increases its own numbers and well-being to the detriment of others. In competition, all parties involved receive less individual benefit from their environment than they would have if they were free to act without competitors or parasites.2
That Simple Biophysical/Economic Model
Hoek et al., 2016, constructed an experimental laboratory culture3 of two different brewing/baking yeast varieties4 to determine if changes in overall community resource availability alone can account for shifts from mutualist/cooperative behavior to parasitic/competitive behavior. As will be seen, it is fortunate -- for the purpose of using the Hoek et al. findings to examine human culture -- that the basic attributes of the yeast varieties involved in the Hoek experiment qualitatively parallel the different strengths and weaknesses that mark cooperating or competing human groups and individuals.
In the yeast co-culture experiment, the chances for observing mutualism were insured by the selection of two yeast varieties that are able to ‘cross-feed’ each other. One of the two yeast varieties concerned (tryptophan-negative, or Trp-neg), produces and then leaks out (i.e., exports) into the surrounding growth medium the amino acid leucine as a waste product, while the other (leucine-negative, or Leu-neg) leaks (exports) the amino acid tryptophan as a by-product of its metabolic activities. As it happens that the tryptophan producer (Leu-neg) does not internally synthesize enough leucine to maximize its own population growth and maintenance, leucine leaked into the liquid environment shared with the Trp-neg yeast is imported and then used by leucine-deficient Leu-neg. Similarly, as the tryptophan-hungry leucine over-producer (Trp-neg) does not synthesize enough tryptophan to maximize its own population growth and maintenance, it imports tryptophan produced and then exported into solution by the Leu-neg yeast variety. The ability of each yeast variety to synthesize and then export particular amino acids of is, in trade- and market-related economics-speak, the ‘comparative advantage’ of each of the two strains of yeast.
It is important to bear in mind while studying and analyzing the results of the evolution of this two-variety yeast culture that in monoculture the Trp-neg yeasts required at least a 2 micromolar initial concentration of tryptophan to avoid collapsing into extinction.5 In contrast, in monoculture the Leu-neg yeasts needed at least an initial 32 micromolar concentration of leucine to avoid meeting the same fate. These minimum needed amino acid solution strength differences mean that the need for externally sourced leucine of the Leu-neg yeast variety is much more limiting than that for tryptophan of the Trp-neg yeast. This makes the metabolically more ‘fragile’ Leu-neg yeast variety much more sensitive to the hard times of amino acid resource scarcity than its more ‘robust’ co-culture partner that can humbly ‘get by’ on less. Being less dependent on tryptophan exports from Leu-neg, the Trp-neg yeast possesses a singular and more permanent (long run) ‘absolute advantage’ over the Leu-neg yeast variety when amino acid resources are more scarce.
The graph below is a slight reworking of Hoek et al.’s Figure 3 in that a third, dashed set of red and green graph lines is added to the original Figure. These two added dashed lines illustrate the relative advantage and disadvantage of living in a co-culture under conditions of steadily increasing resource availability for each of the two yeast varieties. The additional plot lines were obtained by normalizing (dividing by) Leu-neg and Trp-neg cell concentrations in co-culture against their respective background cell concentrations in monoculture. Consequently, ‘multiplier’ line values on the right Y-axis greater than 1.0 indicate there is advantage from living in co-culture, while multiplier line values less than 1.0 show under what circumstances existing in co-culture are disadvantageous to a given yeast variety.
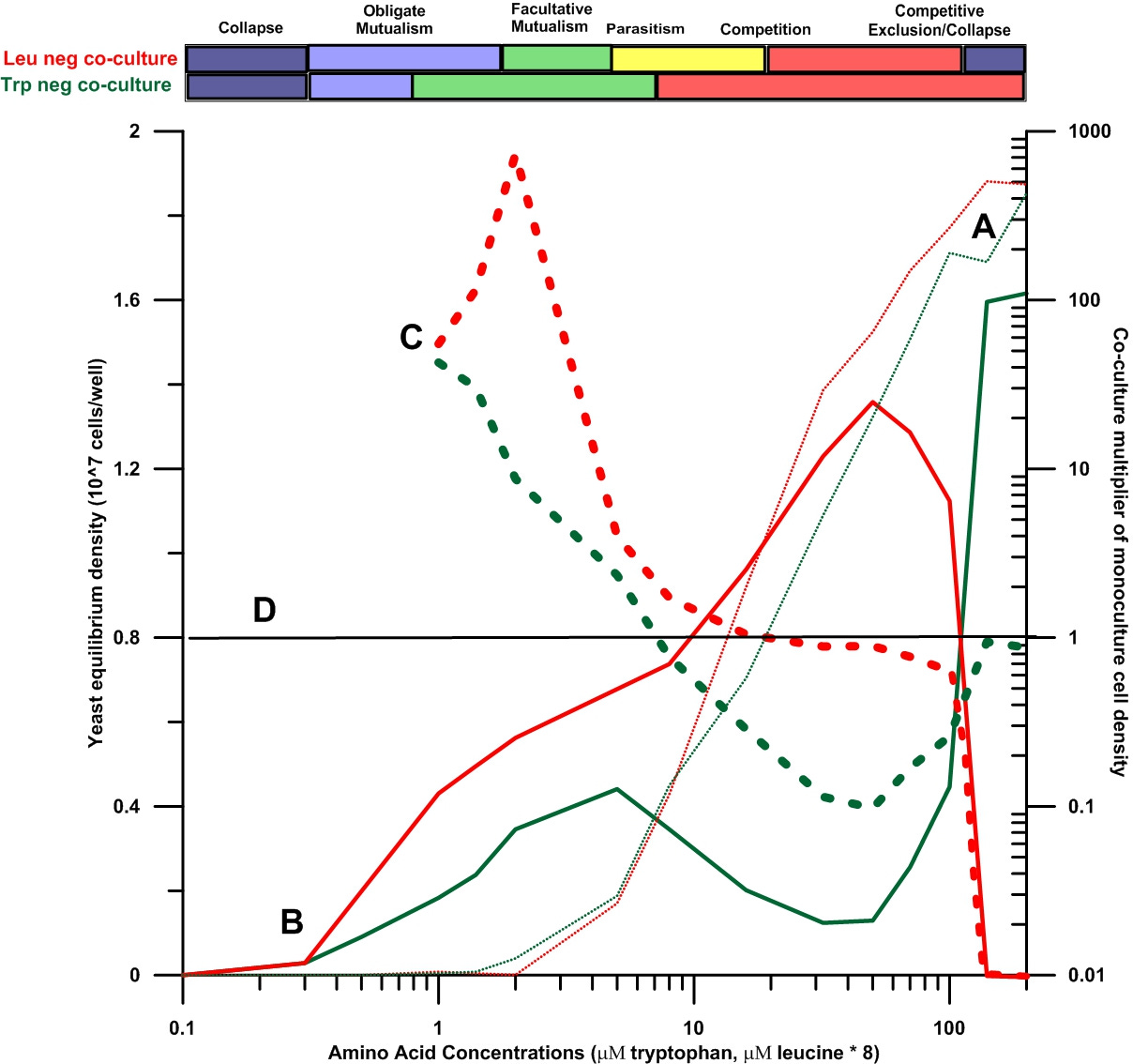
The advantage of adding the red and green dashed lines to the original Hoek graph is that it is easier to comprehend the effects of decreasing and increasing the exogenous concentrations of tryptophan and leucine on the growth of each yeast variety. It also becomes easier to appreciate the different negative and positive effects of yeast growth on each yeast strain in variety isolation or in variety co-culture.
Comparing the two dotted lines labelled at A and the two solid lines labelled at B, it is clear that co-cultured Trp-neg and Leu-neg both maintain much higher population numbers when the growth media is amino acid-poor than is otherwise possible for either yeast variety when they are grown in monocultural isolation under the same impoverished conditions. As already mentioned, this obligate mutualist advantage comes from the facts that in co-culture Trp-neg exports small, but critical, amounts of leucine out to the Leu-neg yeast, and Leu-neg exports smaller, but critical, amounts of tryptophan out to the Trp-neg yeast. Under general amino acid resource scarcity, these small cellular ‘leaks’ of the respective amino acids to the surrounding growth solution are very important to the growth and maintenance of both yeast varieties, increasing the cell numbers of both yeast strains much higher than possible when they dwell in monocultural isolation.
As additional exogenous leucine and tryptophan are added to the growth medium, however, both yeasts can and do become progressively less reliant on each other’s amino acid exports for their cell growth. Obtaining more and more of their needed amino acid requirements abiotically from the externally-enriched environment rather than from the cell leaks of their mutualist partners, the two yeast varieties gradually can afford to become facultative mutualists, obtaining benefit from both their co-culture partners and the outside growth medium. See the courses of the heavy dashed lines labelled at C and note the intermediate range of growth medium amino acid concentrations where the right Y-axis values are greater than 1.0.
The two solid lines labelled B on Figure 1 show that as abiotic, exogenous amino acid concentrations increase, Leu-neg yeast cell numbers increase faster than Trp-neg cell numbers.6 Leu-neg yeast cells are thus – early on -- able to utilize more and more of the environmental resources available to the co-culture as those resources are increased. Leu-neg yeast cells continually sequester most of both critical amino acids to their population – to the material detriment of Trp-neg yeast cells. According to Figure 1, this competitive advantage remains with the Leu-neg yeast variety until abiotic amino acid concentrations and community yeast cell numbers reach quite high levels. Although not explained by Hoek et al., 2016, it is presumed that the observed eventual crash (so-called competitive exclusion by Hoek et al.) of the Leu-neg yeast variety population occurred because of the poisoning by the yeast by-product ethanol produced by the co-cultural community. This poisoning is observed in all conventional brewing operations allowed to proceed to completion, and import of amino acids from growth media into yeast cells has been reported by other researchers to be strongly hindered at high environmental ethanol concentrations. Because Trp-neg yeasts are not nearly as dependent on ethanol poisoning-susceptible amino acid import as Leu-neg yeast cells, this very likely explains recovery of Trp-neg population numbers after the eventual die off/collapse of the Leu-neg yeast population at very high amino acid concentrations.
What might this all have to do with humans?
Despite having many behaviors completely subject to unconscious influences like hormones, age, physical hunger, thirst, and state of health, anthropocentric humans tend to believe their actions and choices are usually under some reliable kind of special cognitive aptitude and control possessed only by homo sapiens. Nevertheless, the Hoek et al. (ibid.) yeast experiment suggests the possibility that human cultural behaviors like cooperation and competition may instead be hardwired as deeply into the basic metabolisms of multi-cellular Man as they are into single-celled yeasts – and, so, also as directly controlled by fluctuations in abiotic and biotic resource abundance.
Is there any recorded historical evidence supporting this possibility?
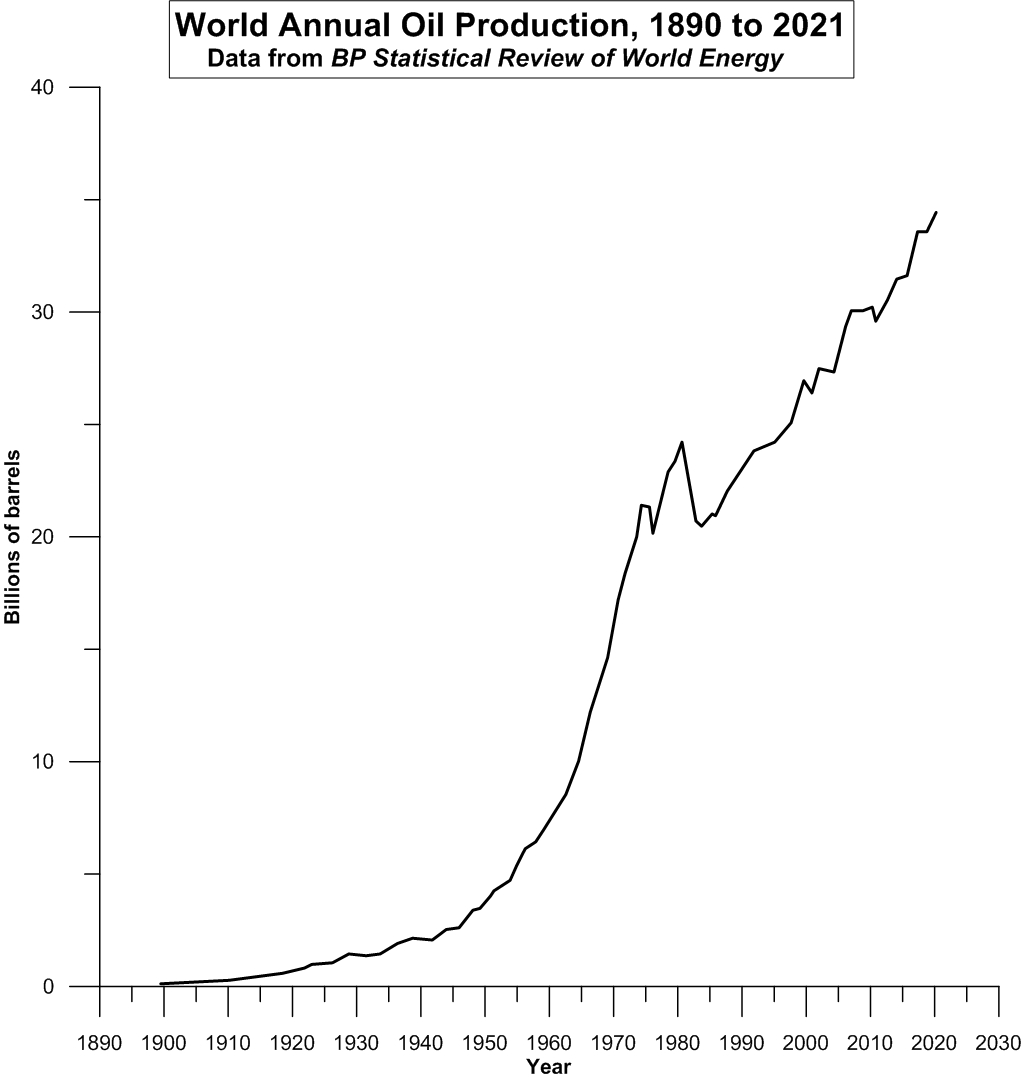
Consider that modern human history parallels the Hoek et al. (ibid.) yeast experiment in that yeast and human populations both benefited numerically from having access to increasing concentrations of abiotic nutrients. In the case of the yeast varieties, their growth medium was progressively enriched in the amino acids, leucine and tryptophan. In the case of humans, on the other hand, sources of “free energy” for conducting all sorts of human economic work have continually increased in abundance since the Industrial Revolution. As an example, the case for that most liquid, most readily obtained and produced, and therefore economically most dominant of such energy sources is illustrated below in Figure 2.
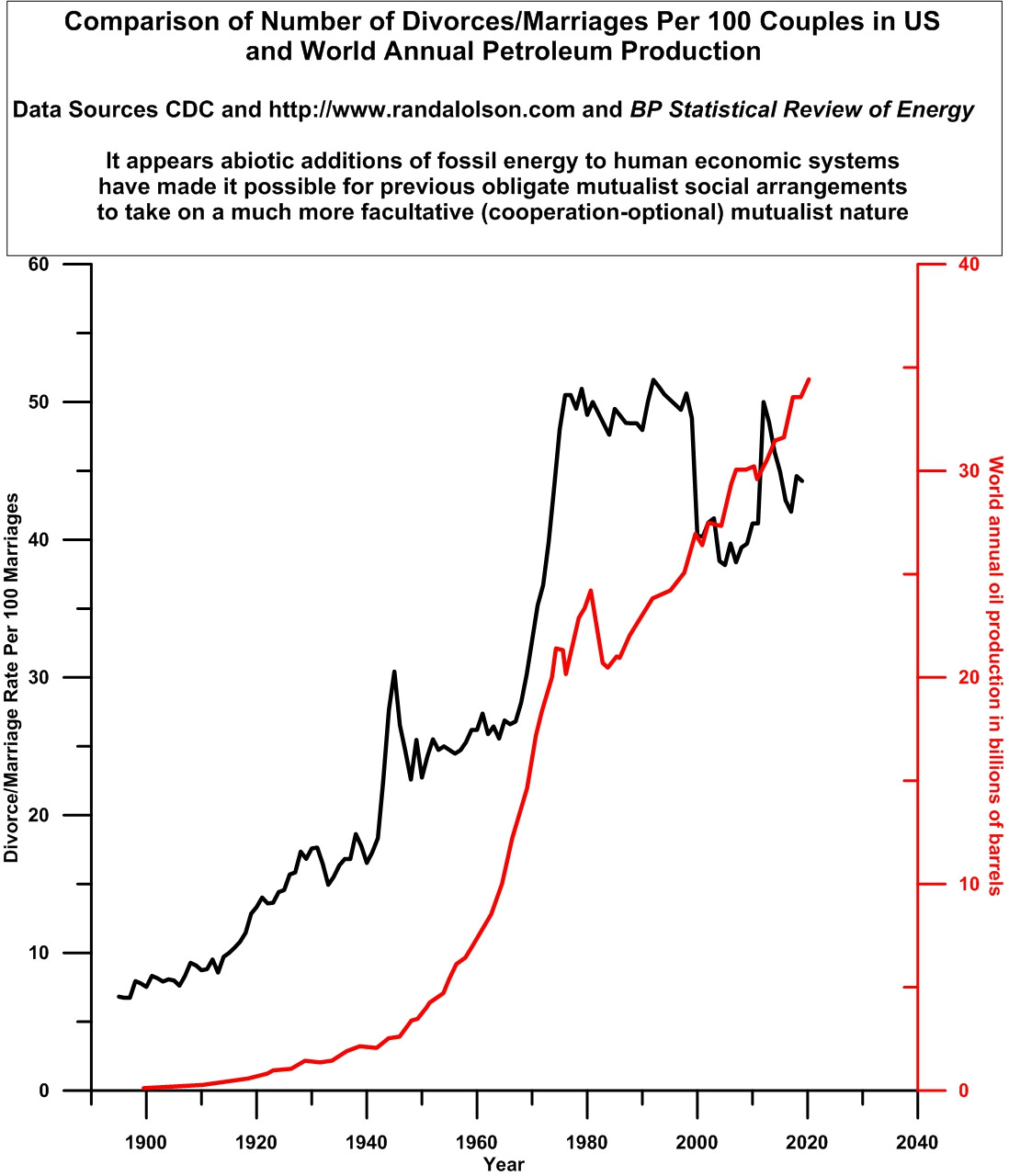
In the yeast case, the while the population of the yeast co-culture became larger as more exogenous tryptophan and leucine were made available, the yeast varieties moved from obligate mutualism at low nutrient conditions to facultative mutualism at intermediate levels of amino acid augmentation. Highest levels of abiotic amino acid enrichment in the yeast experiment eventually pushed both yeast varieties out of formerly beneficial and necessary obligate and facultative mutualism into lose-lose competition and/or win-lose parasitism. In a similar manner, the following graph suggests that the abiotic enrichment of the human growth media with increasing oil (and other fossil fuels) production may have moved the historically traditional and stable, obligate mutualist marriage culture of countries like the US into facultative mutualism where abandonment of the married state became economically feasible for many couples. Currently, of course, many adult females and males in the US (and elsewhere) are in quite fierce competition for jobs in formerly male-dominated professions and industries. See Figure 3.
The yeast experiment showed that different comparative and absolute advantages of the parties engaged in mutualism and competition under conditions of changing resource abundance also affect the size of each mutualist/competitor class. For example, in the yeast experiment, the more opportunistic yeast variety unable to internally produce enough leucine for itself (Leu-neg) was counterintuitively able to increase its numbers faster than the tryptophan-requiring variety (Trp-neg) not quite so reliant on external nutrient inputs. This difference evidently derived from the more opportunistic Leu-neg yeasts’ ability to more rapidly and more efficiently incorporate externally-provided nutrients like leucine. See Figure 1 again.
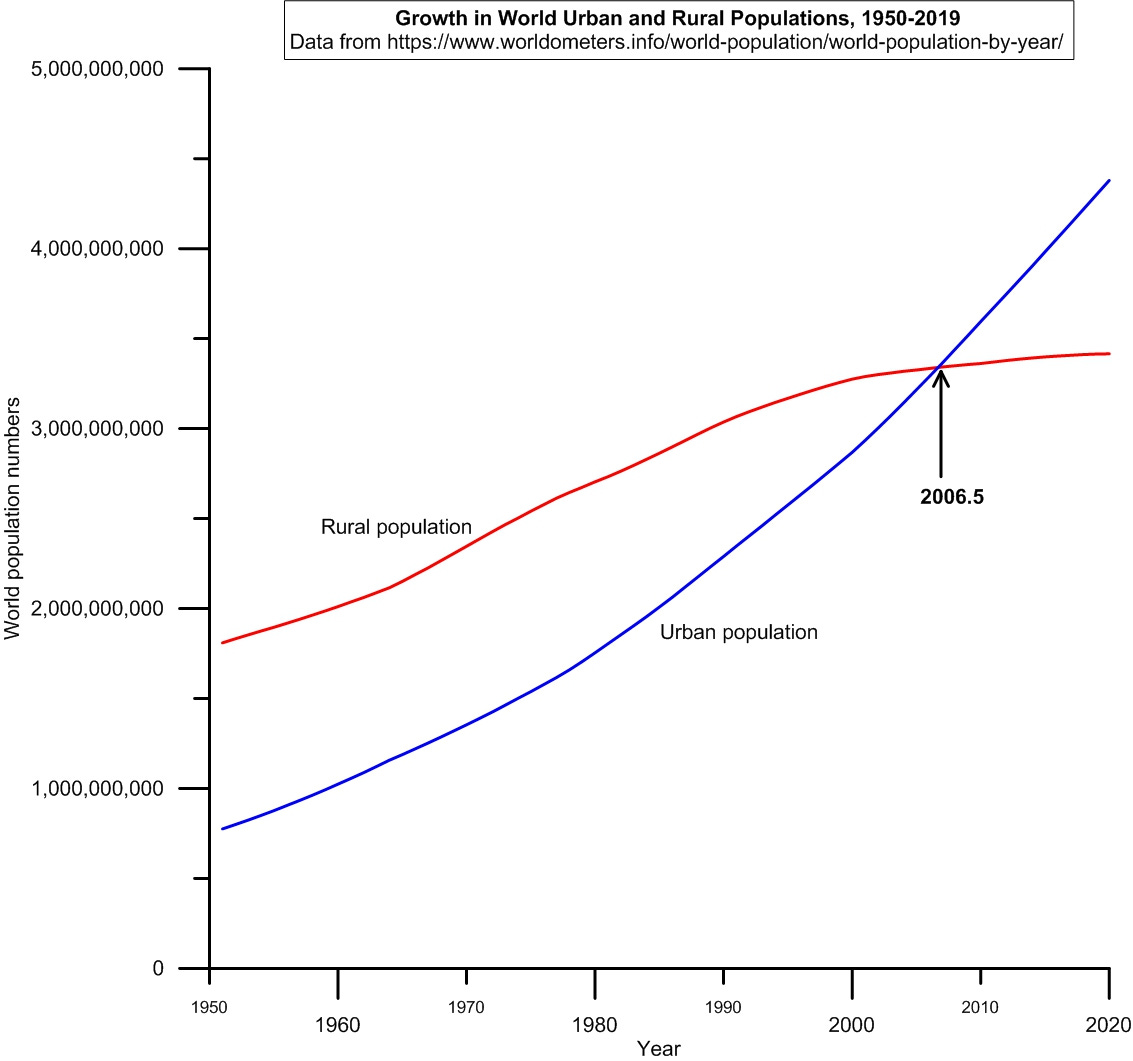
There are, of course, exact correspondences to this yeast co-culture situation in the human world. Since about 3000 BC, humanity has been divided into a co-culture made up of an initially mutualistic rural commodity-producer class and an initially mutualistic urban commodity-consuming class. Not until very recently (2006-2007) has the urban commodity-consuming class outnumbered and dominated the world’s rural commodity-producing population. See Figure 4.
Very interestingly in this regard, in the US the 2006-2007 global changeover to urban population dominance was also marked by shift to urban Blue (Democrat) State median household income dominance over residents of the rural (Republican) Red States. See Figure 5.
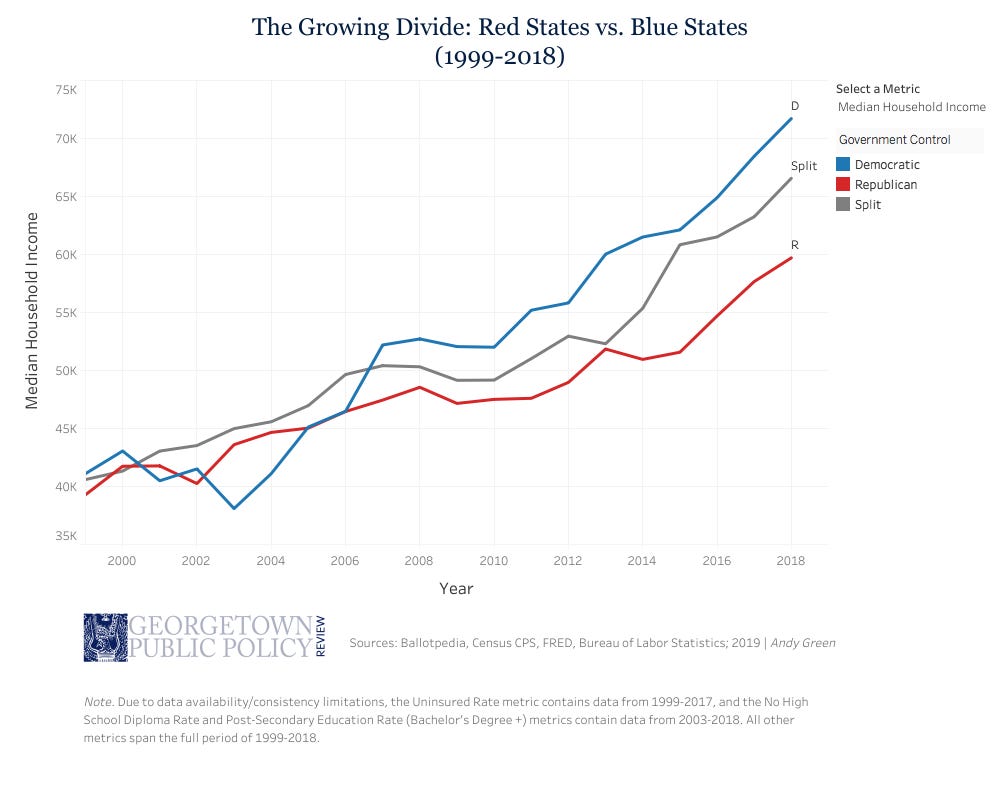
Further, this same transition is also reflected in the distribution of US wealth ownership as illustrated in Figure 6. In the same 2006-2007 time period as urban populations began to dominate in the world population and Blue State occupants’ income began exceeding that of the occupants of Red States, the [urban] top 0.1% ownership share of US wealth began to exceed that wealth possessed by the bottom 80% of US citizens. In the yeast-speak of Figure 1, this shift in wealth transfer signifies that pronounced biological parasitism of rurally-oriented populations and industries has been continuously taking place in the US since this economic and cultural milestone.

Conclusion
Recall that in the Hoek et al. (2016) yeast experiment the Leu-neg yeast variety achieved and maintained numerical and environmental dominance over its Trp-neg former leucine supplier as the shared living environment became more and more enriched in abiotic, externally-provided nutrients. Eventually, however, the absolute advantage of the dominant Leu-neg yeast – i.e., its greater ability to import and utilize (i.e., consume) abiotic nutrients like leucine – disappeared because of developing external conditions, alcohol poisoning, created within the yeast co-culture. Not having the same weaknesses and strengths as the died-off Leu-neg yeast variety, the formerly subordinate Tr-neg yeast members of the yeast co-culture thereafter was able to completely replace the Leu-neg population in the still highly-enriched growth medium.7
Continuing to follow the findings revealed in the yeast experiment to examine and interpret the current human co-culture, the question then becomes: is the urban commodity-consumer class showing any signs of weakening or slackening in its ongoing dominance of the human biological niche? There are a couple of major lines of evidence firmly answering this question in the affirmative.
First, the entire economic business of people is not different in basic principle from that of yeasts. Unlike yeasts, however, people are able to take in and utilize not only physical nutrients from their outside environment to grow and maintain themselves and their communities, but can also incorporate observations, knowledge, and related practical methods and techniques for more effectively and appropriately obtaining and utilizing those physical resources. While the absolutely biologically necessary -- but comparatively simple -- abilities necessary to consume and digest food, and to make use of other goods and services, remain undisturbed and fully effective in the urban commodity-consumer class, that class’s general ability to effectively internalize and utilize information from the outside environment, and even from their own physical bodies, currently is in appreciable disarray.
In the US example, the most striking of all evidence for the ebbing of the human niche dominance of the urban commodity-consumer class, however, is the numerical die-off of urban commodity-consumer replacement observable in Blue State birth rate and consequent family size. See Figure 7.
In sum, using the simple principles brought to light by the biophysical/economic mutualism-competition yeast model of Hoek et al., 2016, modern human discord and break down of cultural cooperation appear to primarily be a result of the steadily increasing physical abundance made possible by human scientific and technological success. As the human environment became progressively resource-enriched over the last 250+ years, human-human interactions formerly more characterized by cooperative mutualism became replaced with those more characterized by antagonistic parasitism and/or competition. There is evidence, however, that the still numerically dominant urban commodity-consumer class of humanity is suffering from waning reproductive and cognitive effectiveness and may lose its present dominance in the human co-culture either because of the many inadvertent side effects of its idiosyncratic way of doing things, and/or because of reversal of the resource enrichment of the co-culturally inhabited human physical environment.
Judging by a consideration of current and recent human events, and by consideration of the resource potential of the earth’s physical environment, it seems most likely to the writer that an initial loss of the cultural dominance of the urban commodity-consumer class will be followed by a partial reversal of the level of resource abundance (‘the nutrient concentration of human growth medium’) available to humanity. This environmental impoverishment should, according the meaning of Figure 1, eventually promote some measure of general return to cooperative mutualism and concord within and between the urban and rural human co-cultures.
Increased, broader competency therefore can sometimes serve as a substitute for a scarce environmental resource or for a partner that becomes unreliable.
See “Blurred lines between competition and parasitism”: https://epublications.marquette.edu/cgi/viewcontent.cgi?article=1570&context=bio_fac
The two yeast varieties in the experiment operate on the same trophic level -- meaning that both yeasts use the same abiotic nutrients extracted from the solution that they live in to grow and duplicate themselves. Neither yeast variety preys on the other as both occupy exactly the same biological niche (https://biologydictionary.net/niche/).
Both are members of https://en.wikipedia.org/wiki/Saccharomyces_cerevisiae. See https://www.pnas.org/doi/full/10.1073/pnas.1313285111 for more information on the strains used.
The concentrations of the two amino acids after a daily 10X dilution with the growth medium.
Hoek et al., 2016, provide no hypothesis or observations explaining this difference, but it is likely that the energy and resources saved by Leu-neg cells in not having to synthesize much leucine for themselves allows them an early faster growth rate than Trp-neg cells. This difference could explain the origin of the parasitism and competitive advantage demonstrated by Leu-neg cells in the experimental results of Hoek et al.(ibid.) under intermediate and high amino acid concentrations.
This same sort of yeast die-off occurs in all brewing operations. Each brewing yeast variety has an upper limit of alcohol tolerance. Once that limit is reached, the yeast concerned generally forms spores to protect itself from alcohol in the outer environment and becomes metabolically inactive. If more water is added to the brew and enough sugar and other nutrients are present in the diluted brew, these spores will reactivate and begin to actively populate the nutrient solution with yeast cells once again. When using this yeast model to interpret human social behavior, the reversible spore formation ability of yeast varieties with changing environmental conditions is definitely something to keep in mind as far as the matter of employing the yeast experiment results to assess the permanence and changeability of culture in Man. Human tendencies that disappear under changed circumstances can also reappear with further changes in the human and natural environments.