
In More Detailed Praise of Quercetin
Regular consumption of quercetin-rich foods or supplements appears to be a partial but effective solution to much of the human aging problem
The afternoon light; unless you drum on a jug and sing, there will be the lament of old age, which is unfortunate.
— Liu I-ming, 1796

Introduction
So, the results of my first 2023-2024 wintertime literature-exploration expedition indicate that the widespread dietary polyphenol, quercetin (along with its biochemically-related dietary source, rutin), is pretty much the big Swiss Army Knife of nutraceuticals. Laboratory findings accumulated over the last 30-40 years or so indicate that, by eating certain fruits, vegetables, and plant seeds enriched in quercetin, us humans have been – all through our species’ food consumption history – unwittingly self-medicating ourselves to one degree or another against the wide range of the chronic diseases that result from the ravages of oxygen and time.

This holiday season post summarizes what is scientifically known of the specific relationships between quercetin and several especially critical biochemical processes now recognized as affecting human healthspan and longevity. Given these findings and the historical precedents of the quite anomalous healthspan and longevity experienced by several isolated and discrete human populations, all forced by unique natural conditions to regularly consume foods highly enriched in quercetin and rutin in times of want, there appears to be good reason for the rest of us to consider increasing and maintaining intake of foods and supplements containing high amounts of the compound. Multiple likely reasons, in fact, for making such a dietary adjustment, extracted from publications about the Okinawan population type-case, are illustrated just below.


Bad News I. Where and How Things First Go Wrong in the Cell
I previously related how human bodies noticeably start showing signs of aging after about 40 years of life, as production of cellular reactive oxygen (ROS) and nitrogen species (RNS) in the cells starts outpacing the natural capacity of the body to quench excesses of these reactive species before they cause irreparable cellular harm.
As might be expected, research shows that the onset of this system-wide, chronic biological system error is at, or very close to, the beginning of such diverse age-related chronic illnesses as heart failure, arterio- and atherosclerosis, hypertension, kidney disease, COPD, obesity, diabetes, non-alcoholic fatty liver disease, intervertebral disc degeneration, arthritis, Parkinson’s Disease, Alzheimer’s Disease, ALS, memory and muscle loss, dementia, fatigue, chronic sleep disturbances, memory and muscle loss, dementia, fatigue, chronic sleep disturbances, and age-related depression.1
Childs et al., 2017 points to the likely upstream ‘geographic’ origin of this lamentably broad river of consequences:
“In his 1881 essay ‘The Duration of Life’, evolutionary biologist A. Weismann made an assertion considered radical in his day. “Death takes place because a worn-out tissue cannot forever renew itself, and because a capacity for increase by means of cell division is not ever-lasting but finite”. Weismann’s far-reaching idea — that an inherent limit to cell division contributed to ageing — lay dormant for more than 80 years until L. Hayflick’s work in 1961 demonstrated that mammalian cells do indeed have a finite capacity for cell division, which is a concept now referred to as the ‘Hayflick Limit’.
Weismann and Hayflick both argued that tissues age because non-dividing cells within that tissue lose the ability to participate in repair. What could not be appreciated at the time was that a class of non-dividing cells induced by cellular stress, now referred to as ‘senescent cells’ (SNCs), are also harmful for an entirely different reason. In addition to not being able to contribute to tissue repair through proliferation, SNCs disrupt normal tissue function by secreting factors that recruit inflammatory cells, remodel the extracellular matrix, trigger unwanted cell death, induce fibrosis, and inhibit stem cell function. SNCs that accumulate over time actively damage the tissues in which they reside and can be directly linked to features of natural ageing.”
Although human cells start falling behind in the work of sufficiently dealing with biochemical stresses at around the age of 40 by conducting needed tissue repair and cellular replacement, problematic senescent cells formed and accumulated in response to these stresses usually do not exist in sufficient quantities to cause obvious and debilitating chronic illnesses in the majority of people until they reach their 60s and 70s.
Senescent cells of the senescence-associated secretory phenotype (SASP) spread their harmful influence to surrounding cells by means of small messenger proteins called cytokines. In the diagram below, the small colored dots represent spreading cytokines secreted and then distributed from the senescent cell ‘seeds’ to other cells.

The interacting major agents inducing aging-related cell senescence are diagram-listed below:

On the other hand, inhibitors of this transformation from healthy proliferating and tissue-repairing cell to harmful senescent cell include:

The exogenous chemical agents, senolytics, already demonstrated to be capable of actually killing off and removing trouble-making senescent cells after they have already been formed and have started spreading their noxious influence, are represented below. Dasatinib is a toxic chemotherapy drug, while the other two named compounds are polyphenols widely distributed in the plant kingdom. Compared to dasatinib, quercetin is innocuous, while fisetin concentrations in plants are extremely low compared to quercetin concentrations:

Again, note the presence of plant-derived quercetin in both of the last two illustrations. As will be discussed further, quercetin removes senescent cells secreting problematic substances causing and spreading cellular inflammation. Moreover, this polyphenol also materially inhibits the chemical actions of several enzymes that are now recognized to accelerate and support the formation and development of numerous chronic illnesses. Quercetin also acts directly and less critically as an ordinary strong antioxidant towards reactive oxygen and nitrogen species.


Bad News II. Secondary Effects of Senescent Cells of the SASP Type
The secretions of senescent cells of the SASP type cause deleterious changes in numerous chemical systems supporting mammalian life. One of the most prominent and most damaging changes induced by SASP secretion into aging tissues and organs is the upregulation of the activity of the CD38 enzyme. As previously related here:
“NAD (nicotinamide adenine dinucleotide) is a very abundant and an absolutely biochemically critical coenzyme derived from the various chemical forms of vitamin B3 and the amino acid tryptophan that, among other things, shuttles electrons around the cytosol and mitochondria to produce all of the energy used by human (and other animal) cells, tissues, and organs. NAD accepts and donates electrons with almost equal facility, so functions readily as both an oxidizing agent and reducing agent. During this electron-shuttling work per se, no NAD is consumed and lost from further energy-generating use by the body – the NAD is just repeatedly reduced and oxidized as each human cell converts food sources to the energy form (ATP) it is uses for metabolism.
CD38 (cyclic ADP-ribose hydrolase) consumes especially large quantities of NAD. This enzyme molecule penetrates through the cell and nuclear membranes of macrophages, the cells of the heart, skeletal muscles, nerves, brain, pulmonary arteries, lymphoid system, lungs, digestive tract, and pancreas. Besides functioning as an activator of extracellular immune cells, CD38 also produces messenger molecules that cause cellular release of calcium – and production of much waste NAM [nicotinamide]. Tellingly, the CD38 enzyme becomes a more and more ubiquitous and active NAD consumer and producer of waste NAM in the cells of animal bodies as they [the animals] age. It is suspected by most researchers in the field that the voracious NAD-consuming activity of CD38 is a major, if not the major, reason for declining NAD concentrations [and declining overall energy levels] observed in aging human bodies.”
Unfortunately, the drop in cellular NAD concentration caused by the increase in chemical activity of CD38, because of the accumulation of senescent cells of the senescence-associated secretory phenotype, has even more consequences. NAD shortages caused by CD38 reduce ‘PARP’ enzyme chemical activity too, causing damaged DNA to go unrepaired -- and even more senescent cells of the SASP type to be created as the programmed cell death and removal of malfunctioning cells becomes no longer a ‘when all else fails’ tissue repair option (ibid.):
“The PARP (poly ADP-ribose polymerase) family of enzymes is involved in the repair of nuclear DNA, genomic stability, and programmed cell death. PARPS use NAD as a substrate during their enzyme activities and produce nicotinamide (NAM) from that NAD as a metabolic waste product.”

Through its greedy consumption of cellular NAD resources and its generation of more waste NAM, upregulation of CD38 activity in cells simultaneously also downregulates the chemical activity of the sirtuin class of enzymes which normally act to repair DNA and prompt cells to quench cell component oxidation by reactive oxygen and nitrogen species. Downregulation of sirtuins by CD38 formed under the direct influence of senescent cells therefore iteratively increases the rate of creation of more and more SASP senescent cells, greater NAD shortages, in a repeating vicious cycle. And so it goes, and goes, and goes, and goes.
It gets even worse, however. The downstream negative effects of increased CD38 enzyme activity, declining NAD levels, and reduced PARP and SIRT enzyme activity, do not stop there. The general state of inflammation induced by cell component and process damage described above that comes with aging, formally termed “inflammaging”, makes the whole sympathetic nervous system (SNS) of the body overactive. And, among many other body-wide effects (like hypertension), the development of an overactive SNS leads to systemic upregulation and increased production of the monoamine oxidase (MAO) family of enzymes. MAO enzymes break down (oxidize) such very important neurotransmitters as dopamine, norepinephrine, and serotonin – and simultaneously release high levels of cell-damaging, reactive hydrogen peroxide (H2O2), ammonia, and aldehydes in particularly critical places of the body like the brain (Alzheimer’s, Parkinson’s, dementia), heart (heart failure), and lungs (COPD). Besides damaging cells and ushering in chronic diseases, the consequent dearth of dopamine, norepinephrine, and 5-HT/serotonin from MAO enzyme chemical activity, would tend to, all other things being equal, make aging people more unhappy, less alert, slower-witted, and more sleep-deprived than when they were younger.
See the review provided by Santin et al., 2021, explaining that this age-related increase in MAO enzymatic activity with its effect on neurotransmitters is ultimately behind the development of such age-related chronic diseases as Parkinson’s, Alzheimer’s, dementia, depression (and age-related anxiety), sleep disturbances, obesity, muscle loss, hearing loss, memory loss, diabetes, cardiac muscle aging and heart failure, gradual loss of lung function, and general fatigue. The three graphics below summarize the text of the review article.



The Primary Effects of Quercetin on Senescent Cells of the SASP Type, and Its Secondary Effects on the CD38 and MAO Enzymes
As shown in the illustration below, scientific research into quercetin is accelerating particularly rapidly following continued discoveries of the positive effects this ubiquitous plant substance on human health, especially with regard to age-related chronic diseases. Speaking of plant polyphenols like quercetin generally, Zamora-Ros et al., 2018, state:
“Over the last two decades, the literature on polyphenols has grown exponentially following the recognition of their antioxidant, anti-inflammatory, and anti-carcinogenic properties and more evidence for their potential beneficial effects upon health.”

Discussed previously, it has been demonstrated in cell culture, animal model, and in limited human clinical studies, that quercetin is capable of removing senescent cells of the SASP type. In vitro cell data indicate that even a very small concentration of quercetin will incrementally reduce the senescent cell burden afflicting humans. From just a little greater than nil µmoles/liter up to 10 µmoles/liter, senescent human umbilical vein endothelial cells (“HUVEC”), susceptible to quercetin, die off without much counterproductive depression of the repair and growth of normal, non-senescent cells. At higher than 10 µmoles/liter quercetin concentrations, however, quercetin increasingly interferes with the proliferation of normal and healthy HUVEC cells. See the graph below for depiction of the above-described cell states.

In slight contrast to these laboratory observations of very young endothelial cells, test of quercetin’s effect on adult human arterial vascular endothelial cells indicates that quercetin begins to kill senescent adult endothelial cells only at about the higher quercetin concentration of 4 µmoles/liter, and begins significantly depressing the activity non-senescent proliferating cells at quercetin concentrations of greater than 8 µmoles/liter.1
In other in vitro laboratory cell tests, it has been shown that quercetin significantly inhibits the chemical activity of the CD38 enzyme, that enzyme which unhealthily creates NAD shortages in aging people, and then multiplies that negative effect by downregulating the expression of the sirt-1 gene and thereby decreasing the ability of body cells to cope with normal ROS and RNS oxidation processes. Escande et al., 2013, demonstrated dose-dependent reduction in CD38 activity at quercetin concentrations starting as low as 0.5 μM/L. See graphs below. A 50% reversible reduction of CD38 enzyme chemical activity was achieved in the laboratory at quercetin concentrations of about 15 μM/L, while that CD38 activity reduction very significantly increased cellular NAD+ concentrations in a quercetin dose-dependent manner.


Turning now to the age-problematic MAO enzymes, recall from the Santin et al., 2021, reference given above, that the development of age-related depression, obesity, Parkinson’s Disease, heart failure, pulmonary disease, metabolic syndrome and diabetes, and various cancers, all appear to be related to the age-dependent increase in the chemical activity of the MAO-A form of the enzyme. Recall also, from the same reference, that age-related MAO-B enzyme upregulation evidently is behind the development of not only Parkinson’s Disease but also Alzheimer’s Disease.
While quercetin has been shown to significantly and helpfully inhibit MAO-A at low absolute quercetin concentrations(50% reduction at ‘noisy’ published literature values averaging ~7.5 μM/L quercetin, but varying – depending on the lab concerned -- from 0.01 to 18 μM/L quercetin), the B form of the MAO enzyme likely requires somewhat higher concentrations of quercetin to achieve 50% reversible inhibition of MAO-B. The again ‘noisy’ published literature values for 50% MAO-B inhibition vary from 5.2 μM/L to 90 μM/L quercetin and average 31.5 μM/L. In addition to Carradori et al., 2014, see also Larit et al., 2018 for references to these widely differing lab results concerning level of quercetin needed for 50% MAO-B and MAO-A inhibition.
Can an Apple (or an Onion) a Day Materially Improve the Nature of the Individual Human Aging Process?
As explained above, laboratory data show that quercetin operates at a broad range of concentration levels against some of the major biochemical mechanisms behind the development of aging-related chronic diseases.
Senescent cells of the SASP type are sensitive to any quercetin concentration above 0 μM/L, while the CD38 enzyme starts being inhibited at quercetin levels as low as 0.50 μM/L and reaches 50% inhibition at ~14 μM/L quercetin. MAO-A activity is reduced by about 50% at a published average of ~7.5 μM/L quercetin concentrations, but 50% MAO-B inhibition by quercetin, according to published work, only occurs once plasma quercetin concentrations have gotten closer to a published average value of ~31.5 μM/L quercetin.
Generally speaking, effectiveness of quercetin at each of these tasks increases with increasing quercetin concentration. However, beyond about the 8 to 10 μM/L quercetin plasma level, quercetin has been observed in the laboratory to begin to significantly interfere with normal cell proliferation, making its presence in human cytoplasm counter-productive. See Post Figure 12 again.
Given the uncertainty regarding the degree of MAO-A and MAO-B enzyme inhibition by the concentration of plasma quercetin, and given the ‘normal cell’ toxicity of quercetin levels higher than 8 to 10 μM/L, it appears from current laboratory results that optimal human benefit from quercetin ingestion is safely reached when plasma levels of quercetin approach, but do not exceed, that 8 to 10 μM/L limit.
With regard to potential dietary application of these laboratory findings, questions that logically come to mind are:
1) What is a normal plasma quercetin level for those eating an ordinary modern diet? Does this level of plasma quercetin, given in vitro laboratory results, likely provide any protection against the development of aging-related chronic diseases?
2) What is the immediate, short-term effect on plasma quercetin levels when a person eats a food relatively rich in quercetin?
3) What is the long-term effect on plasma quercetin levels when a person consistently, day after day, consumes foods containing relatively high levels of quercetin?
4) Does short-term or long-term ingestion of foods rich in quercetin bring a person’s plasma quercetin levels up to the point where the number of senescent cells of the SASP type will be significantly reduced, and – at that level of plasma quercetin -- will the deleterious chemical activity of the age-upregulated CD38 and the two MAO enzymes also be significantly reduced?
5) And, as a check of the modern lab work, were the long-term quercetin plasma values in people historically consuming famine foods that are extraordinarily rich in quercetin like sweet potato, acorn flour, or bitter buckwheat, likely high enough to feasibly explain their characteristic yet unique extended longevity and healthspan?
In regard to the first question, Egert et al., 2008, found baseline plasma quercetin levels averaged about 0.054 μM/L in 36 healthy Germans (primarily university students). Erlund et al., 2000, found similar average levels of about 0.066 μM/L quercetin in the blood of 16 university-aged Finns. Hollman et al., 1997, measured an average 0.017 μM/L quercetin in the plasma of 5 women and 4 men of college age temporarily following a strictly quercetin-free diet. While these low, background plasma quercetin levels should at least marginally help reduce senescent cell populations of the SASP type, inhibition of CD38, and MAO enzyme activity associated with the late life development of serious chronic diseases would have been theoretically minimal to non-existent. Continued into middle-age, the quercetin content of the diet of each of the students concerned would have provided little, if any, potential protection against the development of such diseases.
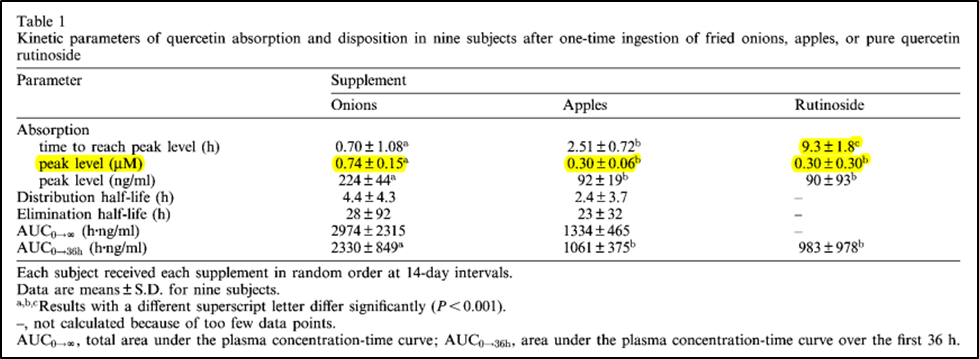
Fortunately, with regard to question #2, ingestion of quercetin-containing foods can change plasma quercetin levels quite markedly. Post Table I below from Hollman et al., 1997, shows peak human plasma levels of quercetin following one-time ingestion of onion or apple containing quercetin, or a supplement of purified rutin. Note that peak plasma quercetin levels are reached relatively rapidly for both onion and apple (0.7 to 2.5 hours), but extremely slowly for rutin (9.3 hours). (Recall here that quercetin is quickly absorbed in the upper gastrointestinal tract, while quercetin-containing rutin is not absorbed and transformed into quercetin until the rutin-containing foods later reach the colon.)
A single meal portion of fried onions, a raw apple, or 200 mg of a rutin supplement all raised average peak plasma quercetin levels well above the 0.017 to 0.066 μM/L values seen in people eating their culturally normal diets. Nevertheless, these single-meal quercetin spikes in plasma quercetin remain below the laboratory-defined thresholds needed for appreciable inhibition of the CD38 and MAO enzymes exacerbating the human aging process.
Fortunately, human data show that quercetin has a long half-life of about 16 hours in the human body. This long half-life means that half of the quercetin added to human plasma on one day is still there about 16 hours later, waiting to be added to the new day’s dietary consumption of the substance. This time-additive nature of regularly consumed dietary quercetin answers questions #3 and #4 above, showing that regular consumption of foods containing notable amounts of quercetin will lead to a gradual increase in plasma quercetin levels, levels closer to the quercetin laboratory thresholds marking beneficial CD38 and MAO enzyme inhibition. For dietary reference, descending average content of quercetin in foods is listed in Post Table II below.

Two studies have examined the cumulative effect of serial, meal-by-meal, day-by-day supplementation of oral quercetin on plasma quercetin levels. The graph below, Post Figure 14, illustrates the combined findings of the two research groups, findings that permit at least rough estimation of the daily quercetin consumption required to reach and then maintain plasma quercetin levels of the lab-indicated magnitude that beneficially removes senescent cells of the SASP type, and reversibly inhibits some of the chemical activity of the harmful age-upregulated CD38 and MAO enzymes.

Quercetin-Rich Famine Foods, Healthspan, and Longevity
Anecdotally, the answer to question #5 above is a qualified yes. Up until relatively recently, two very different, geographically-isolated cultures2 inarguably known and recognized for their characteristically extreme longevity and long healthspan have had to rely on “famine foods” containing unusually high concentrations of quercetin and/or rutin for their staple nutrition. Both of these populations – the island-dwelling Okinawans, and the mountain-dwelling Sardinians – had no choice but to regularly ingest anomalously high amounts of quercetin because of the extreme physical conditions of their living environment. In order to compensate for the effect of typhoon-related flooding on rice culture, the centenarian cohort living at the last turn-of-the-century in Okinawa had to rely on quercetin-rich sweet potato for most of its daily calories until a decade or two after WWII. In order to compensate for the high elevations and steep slopes preventing much grain agriculture, the mountain-dwelling people of Sardinia were formerly very reliant on a staple bread made of rutin-rich acorn flour over the same 1900s time period.

It can be seen from Post Figure 14 that the estimated average plasma quercetin levels of the Okinawans and the Burusho/Hunza would have had appreciable beneficial effect on senescent cells of the SASP type and on control of energy level-reducing and cell repair-reducing CD38 enzyme activity. Because of the current uncertainty regarding quercetin’s power as a MAO enzyme inhibitor, whether or not upregulated MAO-A and MAO-B enzyme activity leading to the development of neurodegenerative conditions in old age would have been significantly diminished by the traditional Okinawan and Burusho famine food diets is not yet clear, however.3
A final note regarding the usual female advantage towards absorbing dietary quercetin because of that sex’s greater apparent ability to transform dietary rutin into quercetin. Unusually, the ratio of mountain Sardinian male centenarians to mountain Sardinian female centenarians is approximately 1:1 – and not the usual 1:3 or less seen elsewhere in the world (including in Okinawa). A possible explanation for this odd longevity ‘equalization’ between the sexes in Sardinia may lie in the long, low-temperature4 cooking process traditionally used in making Sardinian acorn flour bread. Low bitter buckwheat processing temperatures have been shown to permit, for example, the very high levels of rutin in that variety of buckwheat flour to be thoroughly transformed into bitter-tasting quercetin before final, higher temperature bread baking. Such a chemical transformation of rutin into quercetin would have removed any dietary advantage – and presumably augmented longevity and healthspan – that the Sardinian women would have otherwise enjoyed over Sardinian men.
https://sci-hub.ru/10.1016/j.arr.2021.101256, https://www.sciencedirect.com/science/article/pii/S155041312030190X, https://www.sciencedirect.com/science/article/am/pii/S0006322316000809, https://www.nature.com/articles/s41392-023-01577-3, https://www.cell.com/molecular-cell/pdf/S1097-2765(20)30236-7.pdf, https://www.cell.com/cell-metabolism/pdf/S1550-4131(16)30224-8.pdf, etc.
The Burusho/Hunza, with their traditional high-rutin famine food of bitter buckwheat are not mentioned here because of the continuing controversy regarding the verification of the frequency of centenarians and supercentenarians in that subpopulation.
For a n=1 case: my own experimentation with regular quercetin and rutin supplementation has extinguished the (since my 40s) insomnia I usually experience in the fall and winter months. This personal experience indicates that appreciable quercetin inhibition of MAO-A activity in the brain towards the serotonin/5-HT neurotransmitter controlling sleep is taking place despite academic uncertainty about the potency of quercetin as a MAO enzyme inhibitor.
The high altitudes the mountain Sardinians and Burusho live at would have placed an air pressure-dependent limit on the boiling/baking temperatures of acorn and buckwheat flour. Water boils at lower temperatures at high altitudes.